



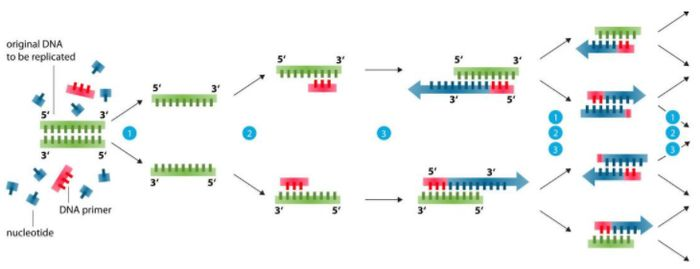
Pohjamaalisuunnittelun perusta (99 % ongelmista voidaan ratkaista)
1. Alukkeen pituus: Oppikirja vaatii 15-30 bp, yleensä noin 20 bp.Todellinen kunto on parempi olla 18-24 bp spesifisyyden varmistamiseksi, mutta mitä pidempi sen parempi, liian pitkä aluke myös heikentää spesifisyyttä ja vähentää saantoa.
2. Alukkeen monistusalue: 200-500 bp on sopiva, ja fragmentti voidaan laajentaa 10 kb:iin tietyissä olosuhteissa.
3. Pohjapohja: G+C:n pitoisuuden tulisi olla 40-60 %, liian pieni G+C:n vahvistusvaikutus ei ole hyvä, liian paljon G+C:tä syntyy helposti epäspesifisiä vyöhykkeitä.ATGC jakautuu parhaiten satunnaisesti välttäen yli 5 puriini- tai pyrimidiininukleotidin klustereita.Multi-gc 5'-päälle ja välisekvensseille stabiilisuuden lisäämiseksi, vältä rikasta GC:tä 3'-päässä, ei GC:tä viimeisille kolmelle emäkselle tai ei GC:tä kolmelle viimeisestä viidestä emäksestä.
4. Vältä toissijaista rakennetta alukkeissa ja vältä komplementaatiota kahden alukkeen välillä, erityisesti komplementaatiota 3'-päässä, muuten muodostuu alukedimeeriä ja syntyy epäspesifisiä monistettuja vyöhykkeitä.
5. Alukkeiden 3 '-pään emäkset, erityisesti viimeinen ja toiseksi viimeinen emäs, tulee yhdistää tiukasti, jotta vältetään parittamattomien terminaalien emästen aiheuttama PCR-virhe.
6. Alukkeissa on tai voidaan lisätä sopivia katkaisukohtia, ja monistetussa kohdesekvenssissä tulisi edullisesti olla sopivat katkaisukohdat, mikä on erittäin hyödyllistä katkaisuanalyysissä tai molekyylikloonauksessa.
7. Alukkeiden spesifisyys: alukkeilla ei pitäisi olla ilmeistä homologiaa muiden nukleiinihapposekvenssitietokannan sekvenssien kanssa.
8. Opi käyttämään ohjelmistoja: PP5, Oligo6, DNAstar, Vector NTI, primer3 (Tämä verkkosuunnittelu toimii parhaiten).
Yllä oleva sisältö voi ratkaista vähintään 99 % pohjamaalin suunnitteluongelmista.
Hallitse pohjamaalin suunnittelun yksityiskohtia
1. Pohjamaalin pituus
Yleinen alukkeen pituus on 18-30 emästä.Yleisesti ottaen tärkein alukkeen hehkutuslämpötilaa määräävä tekijä on alukkeen pituus.Pohjusteen hehkutuslämpötila valitaan yleensä (Tm-arvo -5 ℃), ja jotkut käyttävät suoraan Tm-arvoa.Seuraavia kaavoja voidaan käyttää karkeasti laskettaessa alukkeiden hehkutuslämpötilaa.
Kun alukkeen pituus on alle 20 bp: [4(G+C)+2(A+T)]-5 ℃
Kun alukkeen pituus on yli 20 bp: 62,3℃+0,41℃(%GC)-500/pituus-5 ℃
Lisäksi monia ohjelmistoja voidaan käyttää myös hehkutuslämpötilan laskemiseen, laskentaperiaate on erilainen, joten joskus lasketussa arvossa voi olla pieni aukko.PCR-reaktioiden optimoimiseksi käytetään lyhyimpiä alukkeita, jotka takaavat vähintään 54 ℃:n pariutumislämpötilat parhaan tehokkuuden ja spesifisyyden saavuttamiseksi.
Kaiken kaikkiaan alukkeen spesifisyys kasvaa kertoimella neljä jokaista lisänukleotidia kohti, joten alukkeen vähimmäispituus useimmissa sovelluksissa on 18 nukleotidia.Alukkeen pituuden yläraja ei ole kovin tärkeä, lähinnä reaktiotehokkuuden kannalta.Entropiasta johtuen mitä pidempi aluke, sitä pienempi nopeus, jolla se pariutuu sitoutuakseen kohde-DNA:han muodostaen stabiilin kaksijuosteisen templaatin DNA-polymeraasin sitoutumista varten.
Käytettäessä ohjelmistoa alukkeiden suunnitteluun, alukkeiden pituus voidaan määrittää vuorotellen TM-arvon perusteella, erityisesti fluoresenssikvantitatiivisen PCR:n alukkeiden osalta TM = 60 ℃ tai vastaavaa tulisi kontrolloida.
2.GC-sisältö
Yleensä G+C:n pitoisuus alukesekvenssissä on 40 % - 60 %, ja alukeparin GC-pitoisuus ja Tm-arvo tulisi koordinoida.Jos alukkeella on vakava A-GC- tai AT-taipumus, sopiva määrä A-, T- tai G- ja C-pyrstöä voidaan lisätä alukkeen 5'-päähän.
3. Hehkutuslämpötila
Hehkutuslämpötilan tulee olla 5 ℃ alhaisempi kuin irrotuslämpötila.Jos alukeemästen lukumäärä on pieni, pariutumislämpötilaa voidaan nostaa sopivasti, mikä voi lisätä PCR:n spesifisyyttä.Jos emästen lukumäärä on suuri, hehkutuslämpötilaa voidaan alentaa sopivasti.Pariutumislämpötilaero alukeparin välillä 4 ℃ ~ 6 ℃ ei vaikuta PCR:n saantoon, mutta ihannetapauksessa alukeparin pariutumislämpötila on sama, joka voi vaihdella välillä 55 ℃ ~ 75 ℃.
4. Vältä vahvistusmallin toissijaisen rakenteen aluetta
On parasta välttää templaatin sekundaarirakennealuetta valittaessa monistettua fragmenttia.Kohdefragmentin vakaa toissijainen rakenne voidaan ennustaa ja arvioida asiaankuuluvalla tietokoneohjelmistolla, mikä on hyödyllistä mallineen valinnassa.Kokeelliset tulokset osoittavat, että paisuminen ei usein onnistu, kun laajennettavan alueen vapaa energia (△G) on alle 58,6lkJ/mol.
5. Yhteensopimattomuus kohde-DNA:n kanssa
Kun monistettu kohde-DNA-sekvenssi on suuri, aluke voi sitoutua useisiin kohde-DNA:n osiin, mikä johtaa useiden vyöhykkeiden ilmestymiseen tulokseen.Tällä kertaa on käytettävä BLAST-ohjelmiston testausta, verkkosivustoa:http://www.ncbi.nlm.nih.gov/BLAST/.Valitse Tasaa kaksi sekvenssiä (bl2seq).
Alukesekvenssien liittäminen vyöhykkeelle 1 ja kohde-DNA-sekvenssit vyöhykkeelle 2 on vaihdettavissa, ja BLAST laskee komplementaariset, antisense- ja muut mahdollisuudet, joten käyttäjien ei tarvitse huomata, ovatko molemmat ketjut sense-ketjuja.Voit myös syöttää GI-numeron, jos tiedät sekvenssin GI-numeron tietokannasta, joten sinun ei tarvitse liittää suurta osaa sekvenssistä.Napsauta lopuksi Kohdista kohdassa 3 nähdäksesi, onko alukkeessa useita homologisia kohtia kohde-DNA:ssa.
6. Primer-liitin
Alukkeen 3 'pää on paikka, jossa laajennus alkaa, joten on tärkeää estää epäsuhtautumiset alkamasta sieltä.3'-pää ei saa olla enempää kuin 3 peräkkäistä G:tä tai C:tä, koska tämä aiheuttaa sen, että aluke laukeaa virheellisesti G+C-rikastussekvenssialueella.3'-pää ei voi muodostaa mitään sekundaarirakennetta, paitsi erityisissä PCR- (AS-PCR) -reaktioissa alukkeen 3'-pää ei voi olla yhteensopimaton.Esimerkiksi, jos koodaava alue monistetaan, alukkeen 3'-päätä ei pitäisi lopettaa kodonin kolmanteen asemaan, koska kodonin kolmas asema on altis degeneroitumiselle, mikä vaikuttaa monistuksen spesifisyyteen ja tehokkuuteen.Kun käytät anneksioalukkeita, katso kodonien käyttötaulukkoa, kiinnitä huomiota biologiseen mieltymykseen, älä käytä anneksioalukkeita 3′-päässä ja käytä suurempaa alukkeiden pitoisuutta (1 uM-3 uM).
7. Alukkeiden toissijainen rakenne
Itse alukkeilla ei saa olla komplementaarisia sekvenssejä, muuten alukkeet itse taittuvat hiusneularakenteiksi ja tämä sekundaarirakenne vaikuttaa alukkeiden ja templaattien sitoutumiseen steerisen esteen vuoksi.Jos käytetään keinotekoista harkintaa, itse alukkeiden jatkuva komplementaarinen emäs ei saisi olla suurempi kuin 3 bp.Kahden alukkeen välillä ei pitäisi olla komplementaarisuutta, erityisesti 3'-pään komplementaarista päällekkäisyyttä tulisi välttää alukedimeerien muodostumisen estämiseksi.Yleensä alukeparin välillä ei pitäisi olla enempää kuin 4 peräkkäistä emäshomologiaa tai komplementaarisuutta.
8. Lisää merkkejä tai lokuksia
5'-päällä on vähän vaikutusta amplifikaatiospesifisyyteen, ja siksi sitä voidaan modifioida vaikuttamatta monistusspesifisyyteen.Alukkeen 5'-pään modifikaatio sisälsi: entsyymirestriktiokohdan lisäämisen;Leimattu biotiini, fluoresenssi, digoksiini, Eu3+ jne. Ota proteiinia sitovat DNA-sekvenssit;Mutaatiokohtien tuominen, mutaatiosekvenssien lisääminen ja puuttuminen sekä promoottorisekvenssien lisääminen jne. Ylimääräiset emäkset vaikuttavat enemmän tai vähemmän monistamisen tehokkuuteen ja lisäävät alukedimeerin muodostumisen mahdollisuutta, mutta seuraavaa vaihetta varten on tehtävä joitakin myönnytyksiä.Muita sekvenssejä, joita ei ole kohdesekvenssissä, kuten restriktiokohdat ja promoottorisekvenssit, voidaan lisätä alukkeen 5'-päähän vaikuttamatta spesifisyyteen.Näitä sekvenssejä ei sisällytetä alukkeen Tm-arvojen laskemiseen, mutta niiden komplementaarisuus ja sisäinen sekundaarirakenne tulisi testata.
9. Subkloonit
Suurimman osan ajasta PCR on vain alustavaa kloonausta, ja sitten meidän on subkloonattava kohdefragmentti erilaisiin vektoreihin, joten meidän on suunniteltava lisäemäksiä seuraavaa PCR-vaiheen operaatiota varten.
Alla on yhteenveto joistakin sekvensseistä, jotka on suunniteltu alakloonaukseen.
Restriktioendonukleaasin restriktiokohta lisättiin
Entsyymirestriktiokohtien lisääminen on yleisimmin käytetty menetelmä PCR-tuotteiden alakloonaukseen.Yleensä katkaisukohta on kuusi emästä, katkaisukohdan 5 'pään lisäksi on lisättävä 2-3 suojaavaa emästä.Eri entsyymien tarvitsemien suojaavien emästen määrä on kuitenkin erilainen.Esimerkiksi SalⅠ ei vaadi suojaavaa pohjaa, EcoRⅤ vaatii 1 suojapohjaa, NotⅠ vaatii 2 suojapohjaa ja Hind Ⅲ vaatii 3 suojapohjaa.
LIC lisää hännän
LIC:n koko nimi on Ligation-Independent kloonaus, Navogenin keksimä kloonausmenetelmä erityisesti sen pET-vektorin osaa varten.LIC-menetelmällä valmistetussa pET-kantajassa on ei-komplementaariset 12-15 emäksen yksijuosteiset tahmeat päät, jotka täydentävät vastaavia tahmeita päitä kohdeinserttifragmentissa.Monistusta varten insertoidun fragmentin alukkeen 5'-sekvenssin tulisi täydentää LIC-vektoria.T4-DNA-polymeraasin 3′→5′-ulkopuolinen aktiivisuus voi muodostaa yksijuosteisen tahmean pään insertoituun fragmenttiin lyhyen ajan kuluttua.Koska tuote voidaan muodostaa vain valmistetun inserttifragmentin ja vektorin keskinäisellä pariutumisella, tämä menetelmä on erittäin nopea ja tehokas, ja se on suunnattua kloonausta.
Ohjattu TA-klooni lisää häntä
TA-kloonaus ei kyennyt kohdistamaan fragmenttia vektoriin, joten myöhemmin Invitrogen esitteli vektorin, joka pystyi kohdistamaan kloonaukseen ja joka sisälsi neljä näkyvää GTGGS-emästä toisessa päässä.Siksi PCR-alukkeiden suunnittelussa komplementaarisia sekvenssejä tulisi lisätä vastaavasti, jotta fragmentit voidaan "orientoida".
Jos sinulla ei ole aikaa, voit kokeilla suoraa synteesiä yhdistämällä geenin vektoriin, jota kutsumme musecularists ET-geenisynteesiksi.
D. In-Fusion-kloonausmenetelmä
Ei vaadi ligaasia, ei vaadi pitkää reaktiota.Niin kauan kuin linearisoidun vektorin molemmissa päissä oleva sekvenssi viedään alukkeiden suunnittelussa, sitten PCR-tuote ja linearisoitu vektori lisätään BSA:ta sisältävään infuusioentsyymiliuokseen ja laitetaan huoneenlämpötilaan puoleksi tunniksi, transformaatio voidaan suorittaa.Tämä menetelmä soveltuu erityisen hyvin suuren volyymin muuntamiseen.
10. Yhdistä pohjamaali
Joskus alukkeen suunnittelusta tiedetään vain rajoitettu sekvenssiinformaatio.Esimerkiksi, jos vain aminohapposekvenssi tunnetaan, sulautuva aluke voidaan suunnitella.Sulautumisaluke on seos eri sekvensseistä, jotka edustavat kaikkia eri emäsmahdollisuuksia, jotka koodaavat yhtä aminohappoa.Spesifisyyden lisäämiseksi voit viitata kodonin käyttötaulukkoon vähentääksesi liittämistä eri organismien peruskäyttöasetusten mukaan.Hypoksantiini voidaan yhdistää kaikkien emästen kanssa alentaaksesi pohjusteen hehkutuslämpötilaa.Älä käytä liitettyjä emäksiä alukkeen 3′-päässä, koska viimeisen 3 emäksen pariutuminen 3′-päässä riittää aloittamaan PCR:n väärästä kohdasta.Korkeampia alukepitoisuuksia (1 µM - 3 µM) käytetään, koska alukkeet monissa anneksioseoksissa eivät ole spesifisiä kohdetemplaatille.
PCR-raaka-aineetohjata
1. Pohjusteen määrä
Kunkin alukkeen pitoisuus on 0,1 - 1 umol tai 10 - 100 pmol.Vaadittu tulos on parempi tuottaa pienimmällä pohjamaalimäärällä.Korkea alukkeen pitoisuus aiheuttaa yhteensopimattomuuden ja epäspesifisen monistumisen ja lisää dimeerien muodostumisen mahdollisuutta alukkeiden välille.
2. Pohjustekonsentraatio
Alukkeiden pitoisuus vaikuttaa spesifisyyteen.Optimaalinen alukepitoisuus on yleensä 0,1 - 0,5 μM.Suuremmat alukepitoisuudet johtavat epäspesifisten tuotteiden monistumiseen.
3. Pohjusteen hehkutuslämpötila
Toinen tärkeä parametri alukkeille on sulamislämpötila (Tm).Tämä on lämpötila, kun 50 % alukkeista ja komplementaarisista sekvensseistä on edustettuna kaksijuosteisina DNA-molekyyleina.Tm vaaditaan PCR-pariutuslämpötilan asettamiseen.Ihannetapauksessa pariutumislämpötila on riittävän alhainen varmistamaan alukkeiden tehokkaan pariutumisen kohdesekvenssin kanssa, mutta riittävän korkea vähentämään epäspesifistä sitoutumista.Kohtuullinen hehkutuslämpötila 55 ℃ - 70 ℃.Hehkutuslämpötila asetetaan yleensä 5 ℃ alhaisemmaksi kuin pohjamaalin Tm.
Tm:n asettamiseen on useita kaavoja, jotka vaihtelevat suuresti käytetyn kaavan ja alukkeiden sekvenssin mukaan.Koska useimmat kaavat tarjoavat arvioidun Tm-arvon, kaikki hehkutuslämpötilat ovat vain lähtökohta.Spesifisyyttä voidaan parantaa analysoimalla useita reaktioita, jotka nostavat asteittain pariutumislämpötilaa.Aloita arvioidun Tm-5 ℃:n alapuolelta ja nosta hehkutuslämpötilaa asteittain 2 ℃:n askelin.Korkeampi hehkutuslämpötila vähentää alukedimeerien ja epäspesifisten tuotteiden muodostumista.Parhaan tuloksen saavuttamiseksi kahdella alukkeella tulisi olla likimääräiset Tm-arvot.Jos alukeparien Tm-ero on yli 5 ℃, alukkeet näyttävät merkittävän väärän käynnistyksen käyttämällä syklissä alhaisempaa lämpökäsittelyä.Jos kaksi aluketta Tm ovat erilaiset, aseta hehkutuslämpötila 5 ℃ alhaisemmaksi kuin alhaisin Tm.Vaihtoehtoisesti spesifisyyden lisäämiseksi voidaan suorittaa viisi sykliä ensin korkeammalle Tm:lle suunnitelluissa hehkutuslämpötiloissa, minkä jälkeen loput jaksot hehkutuslämpötiloissa, jotka on suunniteltu pienemmälle Tm:lle.Tämä mahdollistaa osittaisen kopion kohdemallista ahtaissa olosuhteissa.
4. Pohjusteen puhtaus ja stabiilisuus
Mukautettujen alukkeiden standardipuhtaus on riittävä useimpiin PCR-sovelluksiin.Bentsoyyli- ja isobutyyliryhmien poisto suolanpoistolla on minimaalista, eikä se siksi häiritse PCR:ää.Jotkut sovellukset vaativat puhdistusta synteesiprosessin ei-täyspitkien sekvenssien poistamiseksi.Näitä katkaistuja sekvenssejä esiintyy, koska DNA-synteesikemian tehokkuus ei ole 100 %.Tämä on pyöreä prosessi, jossa käytetään toistuvia kemiallisia reaktioita, kun jokainen emäs lisätään DNA:n saamiseksi 3′:sta 5′:aan.Voit epäonnistua kummassakin syklissä.Pidemmät alukkeet, erityisesti yli 50 emästä sisältävät, sisältävät suuren osuuden katkaistuja sekvenssejä, ja ne voivat vaatia puhdistusta.
Alukkeiden saantoon vaikuttavat synteettisen kemian ja puhdistusmenetelmän tehokkuus.Biofarmaseuttiset yritykset, kuten Cytology ja Shengong, käyttävät kaikki vähimmäisOD-yksikköä varmistaakseen oligonukleosidin kokonaistuotannon.Mukautetut pohjamaalit toimitetaan kuivajauheena.On parasta liuottaa alukkeet uudelleen TE:hen niin, että lopullinen pitoisuus on 100 μM.TE on parempi kuin deionisoitu vesi, koska veden pH on usein hapan ja aiheuttaa oligonukleosidien hydrolyysiä.
Pohjusteiden stabiilisuus riippuu varastointiolosuhteista.Kuivajauhe ja liuenneet pohjamaalit tulee säilyttää -20 ℃:ssa.TE:hen liuenneita alukkeita, joiden pitoisuus on suurempi kuin 10 μM, voidaan säilyttää vakaasti -20 ℃:ssa 6 kuukautta, mutta niitä voidaan säilyttää vain huoneenlämmössä (15 ℃ - 30 ℃) alle 1 viikon ajan.Kuivajauhepohjamaaleja voidaan säilyttää -20 C:ssa vähintään 1 vuoden ja huoneenlämmössä (15 C - 30 C) enintään 2 kuukautta.
5. Entsyymit ja niiden pitoisuudet
Tällä hetkellä käytetty Taq-DNA-polymeraasi on pohjimmiltaan koliformisten bakteerien syntetisoima geenitekniikkaentsyymi.Tyypillisen PCR-reaktion katalysoimiseen tarvittava entsyymimäärä on noin 2,5 U (tarkoittaa 100 ul:n kokonaisreaktiotilavuutta).Jos pitoisuus on liian korkea, se voi johtaa epäspesifiseen vahvistumiseen;jos pitoisuus on liian pieni, synteettisen tuotteen määrä vähenee.
6. dNTP:n laatu ja pitoisuus
dNTP:n laatu liittyy läheisesti PCR-monistuksen pitoisuuteen ja tehokkuuteen.DNTP-jauhe on rakeista, ja sen vaihtelevuus menettää biologisen aktiivisuutensa, jos sitä säilytetään väärin.dNTP-liuos on hapan, ja sitä tulisi käyttää korkeina pitoisuuksina, 1 M NaOH:n tai 1 M Tris.HCL-puskuriliuoksen kanssa sen pH:n säätämiseksi arvoon 7,0 ~ 7,5, pieni määrä alipakkauksia, pakastevarastointi -20 ℃.Usein jäädytys-sulatus heikentää dNTP:tä.PCR-reaktiossa dNTP:n tulisi olla 50 - 200 umol/l.Erityisesti tulee kiinnittää huomiota siihen, että neljän DNTPS:n pitoisuuden tulee olla yhtä suuri (saa moolivalmiste).Jos jonkin niistä pitoisuus on erilainen kuin muiden (suurempi tai pienempi), syntyy epäsopivuus.Liian pieni pitoisuus vähentää PCR-tuotteiden saantoa.dNTP voi yhdistyä Mg2+:n kanssa ja vähentää vapaan Mg2+:n pitoisuutta.
7. Templaatti (kohdegeeni) nukleiinihappo
Templaattinukleiinihapon määrä ja puhdistusaste on yksi PCR:n onnistumisen tai epäonnistumisen avainlinkkeistä.Perinteisissä DNA-puhdistusmenetelmissä näytteiden pilkkomiseen ja hävittämiseen käytetään yleensä SDS:ää ja proteaasi K:tä.SDS:n päätehtävät ovat: liuottaa lipidejä ja proteiineja solukalvolla, mikä tuhoaa solukalvon liuottamalla kalvoproteiineja, ja dissosioida tumaproteiineja solussa, SDS voi myös yhdistyä proteiineihin ja saostua;Proteaasi K voi hydrolysoida ja pilkkoa proteiineja, erityisesti DNA:han sitoutuneita histoneita, ja käyttää sitten orgaanista liuotinta fenolia ja kloroformia proteiinien ja muiden solukomponenttien uuttamiseen ja käyttää etanolia tai isopropyylialkoholia nukleiinihapon saostamiseen.Uutettua nukleiinihappoa voidaan käyttää templaattina PCR-reaktioissa.Yleisissä kliinisissä näytteissä voidaan käyttää nopeaa ja yksinkertaista menetelmää solujen liuottamiseen, patogeenien lysoimiseen, proteiinien pilkkomiseen ja poistamiseen kromosomeista vapaiksi kohdegeeneiksi, ja sitä voidaan käyttää suoraan PCR-monistukseen.RNA-templaatin uuttamisessa käytetään yleensä guanidiini-isotiosyanaatti- tai proteaasi K -menetelmää estämään RNaasi hajottamasta RNA:ta.
8.Mg2+-pitoisuus
Mg2+:lla on merkittävä vaikutus PCR-monistuksen spesifisyyteen ja saantoon.Yleisessä PCR-reaktiossa, kun erilaisten dNTP-yhdisteiden pitoisuus on 200 umol/l, sopiva Mg2+-pitoisuus on 1,5 - 2,0 mmol/l.Mg2+-pitoisuus on liian korkea, reaktion spesifisyys laskee, tapahtuu epäspesifistä monistumista, liian pieni pitoisuus vähentää Taq DNA -polymeraasin aktiivisuutta, mikä johtaa reaktiotuotteiden vähenemiseen.
Magnesiumionit vaikuttavat useisiin PCR:n näkökohtiin, kuten DNA-polymeraasiaktiivisuuteen, joka vaikuttaa saantoon;Toinen esimerkki on alukkeen annealing, joka vaikuttaa spesifisyyteen.dNTP ja templaatti sitoutuvat magnesium-ioneihin vähentäen entsyymiaktiivisuuteen tarvittavan vapaan magnesiumionin määrää.Optimaalinen magnesiumionipitoisuus vaihtelee eri alukepareille ja templaateille, mutta tyypillinen PCR-aloituskonsentraatio 200 µM dNTP:llä on 1,5 mM (huomaa: Käytä reaaliaikaisessa kvantitatiivisessa PCR:ssä 3–5 mM magnesiumioniliuosta fluoresoivan anturin kanssa).Vapaiden magnesiumionien korkeammat pitoisuudet lisäävät saantoa, mutta lisäävät myös epäspesifistä vahvistusta ja heikentävät tarkkuutta.Optimaalisen pitoisuuden määrittämiseksi suoritettiin magnesium-ionititraukset 0,5 mM:n välein 1 mM:sta 3 mM:iin.Magnesiumionien optimoinnista riippuvuuden vähentämiseksi voidaan käyttää Platinum Taq DNA -polymeraasia.Platina Taq DNA -polymeraasi pystyy ylläpitämään toimintaa laajemmalla magnesiumionipitoisuuksien alueella kuin Taq DNA -polymeraasi ja vaatii siksi vähemmän optimointia.
9. Pcr:tä edistävät lisäaineet
Hehkutuslämpötilan, alukkeen suunnittelun ja magnesium-ionikonsentraation optimointi riittää useimpien templaattien erittäin spesifiseen monistamiseen;Jotkut mallit, mukaan lukien korkean GC-pitoisuuden omaavat mallit, vaativat kuitenkin lisätoimenpiteitä.DNA:n sulamislämpötilaan vaikuttavat lisäaineet tarjoavat toisen tavan parantaa tuotteen spesifisyyttä ja saantoa.Parhaan tuloksen saavuttamiseksi vaaditaan mallin täydellinen denaturointi.
Lisäksi sekundäärinen rakenne estää alukkeen sitoutumisen ja entsyymin pidentymisen.
PCR-lisäaineet, mukaan lukien formamidi, DMSO, glyseriini, betaiini ja PCRx Enhancer Solution, tehostavat monistumista.Niiden mahdollinen mekanismi on alentaa sulamislämpötilaa, mikä helpottaa alukkeiden pariutumista ja auttaa DNA-polymeraasin pidentymistä sekundaarirakenteen alueen läpi.PCRx-ratkaisulla on muita etuja.Minimaalinen magnesiumionioptimointi vaaditaan käytettäessä Platinum Taq DNA -polymeraasin ja Platinum Pfx DNA -polymeraasin kanssa.Siten Platinum-tekniikka yhdistetään lisäaineeseen spesifisyyden lisäämiseksi ja samalla vähentäen kolmannen lähestymistavan, magnesiumionien optimoinnin, riippuvuutta.Parhaan tuloksen saavuttamiseksi lisäaineiden, erityisesti DMSO:n, formamidin ja glyserolin, pitoisuudet tulee optimoida, sillä ne estävät Taq DNA -polymeraasia.

10. Kuuma aloitus
Hot start PCR on yksi tärkeimmistä menetelmistä parantaa PCR-spesifisyyttä hyvän alukesuunnittelun lisäksi.Vaikka Taq DNA -polymeraasin optimaalinen elongaatiolämpötila on 72 ℃, polymeraasi pysyy aktiivisena huoneenlämpötilassa.Näin ollen epäspesifisiä tuotteita syntyy, kun pitolämpötila on alempi kuin hehkutuslämpötila PCR-reaktion valmistuksen aikana ja lämpösyklin alussa.Kun nämä epäspesifiset tuotteet on muodostettu, ne monistuvat tehokkaasti.Hot-start PCR on erityisen tehokas, kun alukkeiden suunnitteluun käytettyjä kohtia rajoittaa geneettisten elementtien sijainti, kuten kohdistetut mutaatiot, ekspressiokloonaus tai DNA-muokkaukseen käytettyjen geneettisten elementtien rakentaminen ja manipulointi.
Yleinen menetelmä rajoittaa Taq DNA -polymeraasin aktiivisuutta on valmistaa PCR-reaktioliuos jäillä ja sijoittaa se esilämmitettyyn PCR-laitteeseen.Tämä menetelmä on yksinkertainen ja halpa, mutta se ei täydennä entsyymin aktiivisuutta eikä siksi täysin eliminoi epäspesifisten tuotteiden monistumista.
Terminen esikäsittely viivästyttää DNA-synteesiä estämällä olennaista komponenttia, kunnes PCR-laitteisto saavuttaa denaturaatiolämpötilan.Useimmat manuaaliset lämpöaloitusmenetelmät, mukaan lukien Taq DNA -polymeraasin viivästetty lisäys, ovat hankalia, erityisesti suuritehoisissa sovelluksissa.Muissa lämpöpohjustusmenetelmissä käytetään vahasuojaa olennaisen komponentin, mukaan lukien magnesium-ionien tai entsyymien, sulkemiseen tai reaktiivisten komponenttien, kuten templaattien ja puskureiden, fysikaaliseen eristämiseen.Lämpösyklin aikana eri komponentit vapautuvat ja sekoittuvat keskenään vahan sulaessa.Manuaalisen kuumakäynnistysmenetelmän tapaan vahasuojausmenetelmä on hankala ja altis kontaminaatiolle, eikä se sovellu korkean suorituskyvyn sovelluksiin.
Platina-DNA-polymeraasi on kätevä ja tehokas automaattiseen kuumakäynnistys-PCR:ään.Platina Taq DNA -polymeraasi koostuu rekombinantista Taq DNA -polymeraasista yhdistettynä monoklonaaliseen vasta-aineeseen Taq DNA -polymeraasia vastaan.Vasta-aineet formuloidaan PCR:llä inhiboimaan entsyymiaktiivisuutta pitkäaikaisen lämpötilan ylläpitämisen aikana.Taq DNA-polymeraasi vapautui reaktioon denaturaatiovaiheen 94 ℃:n eristyksen aikana, mikä palauttaa täyden polymeraasiaktiivisuuden.Toisin kuin kemiallisesti modifioitu Taq-DNA-polymeraasi lämpöaloitusta varten, platinaentsyymi ei vaadi pitkäkestoista eristystä 94 ℃:ssa (10-15 minuuttia) polymeraasin aktivoimiseksi.PlatinumTaq DNA -polymeraasilla 90 % Taq DNA -polymeraasiaktiivisuudesta palautui 2 minuutin kuluttua 94 ℃:ssa.

Foreasy HS Taq DNA -polymeraasi
11. Nest-PCR
Peräkkäiset monistuskierrokset käyttämällä sisäkkäisiä alukkeita voivat parantaa spesifisyyttä ja herkkyyttä.Ensimmäinen kierros on vakiovahvistus 15-20 sykliä.Pieni osa alkuperäisestä monistustuotteesta laimennettiin 100 - 1000 kertaa ja lisättiin toiselle monistuskierrokselle 15 - 20 sykliksi.Vaihtoehtoisesti alkuperäinen monistettu tuote voidaan mitoittaa geelipuhdistuksella.Toisella monistuskierroksella käytetään sisäkkäistä aluketta, joka voi sitoutua kohdesekvenssiin ensimmäisen alukkeen sisällä.Sisäkkäisen PCR:n käyttö vähentää useiden kohdekohtien monistumisen mahdollisuutta, koska molemmille alukesarjoille komplementaarisia kohdesekvenssejä on vähän.Sama syklien kokonaismäärä (30 - 40) samoilla alukkeilla amplifioi epäspesifiset kohdat.Nested PCR lisää rajoitettujen kohdesekvenssien herkkyyttä (esim. harvinaiset mrnat) ja parantaa vaikean PCRS:n (esim. 5′ RACE) spesifisyyttä.
12. Laskeva PCR
Laskeva PCR parantaa spesifisyyttä käyttämällä tiukkoja pariutumisolosuhteita muutaman ensimmäisen PCR-syklin aikana.Jakso alkaa hehkutuslämpötilasta, joka on noin 5 ℃ korkeampi kuin arvioitu Tm, minkä jälkeen kutakin sykliä alennetaan 1 ℃ - 2 ℃, kunnes hehkutuslämpötila on alle Tm 5 ℃.Vain korkeimman homologian omaava kohdemallipohja monistetaan.Nämä tuotteet jatkavat laajentumistaan myöhemmissä sykleissä syrjäyttäen vahvistetut epäspesifiset tuotteet.Laskeva PCR on käyttökelpoinen menetelmissä, joissa alukkeen ja kohdetemplaatin välistä homologiaa ei tunneta, kuten AFLP DNA-sormenjäljet.
Aiheeseen liittyvät PCR-sarjat
 PCR Easyᵀᴹ (väriaineen kanssa)
PCR Easyᵀᴹ (väriaineen kanssa)
2× PCR-sankariTMSekoitusjärjestelmällä on korkeampi toleranssi PCR-estäjiä kohtaan kuin tavallisella PCR Mix -järjestelmällä, ja se selviytyy helposti useiden monimutkaisten templaattien PCR-monistuksesta.Ainutlaatuinen reaktiojärjestelmä ja tehokas Taq Hero tekevät PCR-reaktiosta korkeamman amplifikaatiotehokkuuden, spesifisyyden ja herkkyyden.
 PCR Heroᵀᴹ (väriaineen kanssa)
PCR Heroᵀᴹ (väriaineen kanssa)
Korkeampi vahvistusteho
Sillä on 5'→3'-DNA-polymeraasiaktiivisuutta ja 5'→3'-eksonukleaasiaktiivisuutta, ilman 3'→5'-eksonukleaasiaktiivisuutta.
Reaaliaikainen PCR Easyᵀᴹ-SYBR Green I Kit
Spesifinen - optimoitu puskuri ja kuumakäynnistys Taq-entsyymi voivat estää epäspesifisen monistumisen ja alukedimeerin muodostumisen
Suuri herkkyys – voi havaita mallin alhaiset kopiot
Pakkaus käyttää ainutlaatuista Foregene-käänteistranskriptioreagenssia ja Foregene HotStar Taq DNA -polymeraasia yhdistettynä ainutlaatuiseen reaktiojärjestelmään, joka parantaa tehokkaasti monistustehokkuutta ja reaktion spesifisyyttä.
Postitusaika: 09-09-2023



